mRNK, tRNK, RRNK - uchta asosiy nuklein kislotalarning o'zaro ta'siri va tuzilishi sitologiya fani tomonidan ko'rib chiqiladi. Bu hujayralardagi transportning (tRNK) rolini aniqlashga yordam beradi. Bu juda kichik, lekin ayni paytda inkor etib bo'lmaydigan muhim molekula tanani tashkil etuvchi oqsillarni birlashtirish jarayonida ishtirok etadi.
tRNKning tuzilishi qanday? Ushbu moddaga "ichkaridan" qarash, uning biokimyosi va biologik rolini bilish juda qiziq. Shuningdek, tRNKning tuzilishi va uning oqsil sintezidagi roli qanday bog'liq?
tRNK nima, u qanday tuzilgan?
Transport ribonuklein kislotasi yangi oqsillarni qurishda ishtirok etadi. Barcha ribonuklein kislotalarning deyarli 10% transportdir. Molekula qanday kimyoviy elementlardan hosil bo'lganligini aniq tushunish uchun tRNKning ikkilamchi strukturasining tuzilishini tasvirlab beramiz. Ikkilamchi tuzilma elementlar orasidagi barcha asosiy kimyoviy aloqalarni hisobga oladi.
Polinukleotid zanjiridan iborat. Undagi azotli asoslar vodorod bog'lari bilan bog'langan. DNK singari, RNK ham 4 ta azotli asosga ega: adenin, sitozin, guanin va urasil. Ushbu birikmalarda adenin har doim urasil bilan, guanin esa odatdagidek sitozin bilan bog'lanadi.
Nima uchun nukleotidda ribo- prefiksi mavjud? Oddiy qilib aytganda, nukleotid bazasida pentoza o'rniga riboza bo'lgan barcha chiziqli polimerlar ribonuklein deb ataladi. Transfer RNK esa aynan shunday ribonuklein polimerning 3 turidan biridir.
tRNKning tuzilishi: biokimyo
Keling, molekula tuzilishining eng chuqur qatlamlarini ko'rib chiqaylik. Ushbu nukleotidlar 3 ta komponentdan iborat:
- Saxaroza, RNKning barcha turlari ribozani o'z ichiga oladi.
- Fosfor kislotasi.
- Azotli va pirimidinlar.

Azotli asoslar bir-biriga kuchli bog'lar orqali bog'langan. Bazalarni purin va pirimidinga bo'lish odatiy holdir.
Purinlar adenin va guanindir. Adenin bir-biriga bog'langan 2 ta halqadan iborat adenil nukleotidiga to'g'ri keladi. Va guanin bir xil "bitta halqali" guanin nukleotidiga to'g'ri keladi.
Piramidinlar sitozin va urasildir. Pirimidinlar bitta halqali tuzilishga ega. RNKda timin yo'q, chunki u urasil kabi element bilan almashtiriladi. Bu tRNKning boshqa strukturaviy xususiyatlariga e'tibor berishdan oldin tushunish kerak.
RNK turlari
Ko'rib turganingizdek, tRNKning tuzilishini qisqacha ta'riflab bo'lmaydi. Molekulaning maqsadi va uning haqiqiy tuzilishini tushunish uchun biokimyoni chuqur o'rganish kerak. Yana qanday ribosoma nukleotidlari ma'lum? Matritsali yoki informatsion va ribosomali nuklein kislotalar ham mavjud. mRNK va RRNK sifatida qisqartiriladi. Tananing to'g'ri tuzilgan oqsil globullarini olishini ta'minlash uchun barcha 3 molekula hujayrada bir-biriga yaqin ishlaydi.

Bitta polimerning ishini 2 ta polimerning yordamisiz tasavvur qilib bo'lmaydi. tRNKning strukturaviy xususiyatlari ribosomalarning ishi bilan bevosita bog'liq bo'lgan funktsiyalar bilan birgalikda ko'rib chiqilsa, yanada aniqroq bo'ladi.
mRNK, tRNK va RRNK tuzilishi ko'p jihatdan o'xshashdir. Ularning barchasida riboza mavjud. Biroq, ularning tuzilishi va funktsiyalari boshqacha.
Nuklein kislotalarning kashfiyoti
Shveytsariyalik Iogan Misher 1868 yilda hujayra yadrosida keyinchalik nukleinlar deb atalgan makromolekulalarni kashf etdi. "Nuklein" nomi (yadro) - yadro so'zidan kelib chiqqan. Biroz vaqt o'tgach, yadroga ega bo'lmagan bir hujayrali mavjudotlarda ham bu moddalar mavjudligi aniqlandi. 20-asrning o'rtalarida u nuklein kislotalar sintezini kashf etgani uchun Nobel mukofotiga sazovor bo'ldi.
oqsil sintezida
Ismning o'zi - transfer RNK - molekulaning asosiy funktsiyasi haqida gapiradi. Bu nuklein kislota o'zi bilan ribosoma RNK tomonidan ma'lum bir oqsil hosil qilish uchun zarur bo'lgan muhim aminokislotalarni "olib keladi".
tRNK molekulasi kam funksiyalarga ega. Birinchisi, mRNK kodonini aniqlash, ikkinchi funktsiya - oqsil sintezi uchun qurilish bloklari - aminokislotalarni etkazib berish. Ba'zi boshqa mutaxassislar qabul qiluvchi funktsiyasini ta'kidlaydilar. Ya'ni, kovalent printsip bo'yicha aminokislotalarning qo'shilishi. Aminosil-TRNA sintataza kabi ferment bu aminokislotalarni "biriktirishga" yordam beradi.
tRNKning tuzilishi uning funktsiyalari bilan qanday bog'liq? Ushbu maxsus ribonuklein kislotasi shunday tuzilganki, bir tomonda doimo juft bo'lib bog'langan azotli asoslar mavjud. Bular bizga ma'lum bo'lgan elementlar - A, U, C, G. Aynan 3 ta "harf" yoki azotli asoslar antikodonni tashkil qiladi - kodon bilan komplementarlik printsipiga ko'ra o'zaro ta'sir qiluvchi elementlarning teskari to'plami.
tRNK strukturasining bu muhim xususiyati shablon nuklein kislotasini dekodlashda xatolik bo'lmasligini ta'minlaydi. Axir, aminokislotalarning aniq ketma-ketligi tanaga hozirda zarur bo'lgan protein to'g'ri sintezlanganligini aniqlaydi.
Strukturaviy xususiyatlar
tRNKning strukturaviy xususiyatlari va uning biologik roli qanday? Bu juda qadimiy tuzilma. Uning o'lchamlari 73 dan 93 gacha nukleotidlar orasida. Moddaning molekulyar og'irligi 25000-30000.
tRNKning ikkilamchi strukturasi tuzilishini molekulaning 5 ta asosiy elementini oʻrganish orqali tahlil qilish mumkin. Demak, bu nuklein kislota quyidagi elementlardan iborat:
- ferment bilan aloqa qilish uchun halqa;
- ribosoma bilan aloqa qilish uchun halqa;
- antikodon halqasi;
- qabul qiluvchi novda;
- antikodonning o'zi.
Va ular ikkilamchi tuzilishda kichik o'zgaruvchan pastadirni ham ajratib turadilar. Barcha turdagi tRNKlarning bir qo'li bir xil - ikkita sitozin qoldig'i va bitta adenozin qoldig'i. Aynan shu nuqtada ulanish mavjud bo'lgan 20 ta aminokislotadan 1 tasi bilan sodir bo'ladi. Har bir aminokislota o'z fermentiga, o'ziga xos aminoatsil-tRNKga ega.

Har bir insonning tuzilishini shifrlaydigan barcha ma'lumotlar DNKning o'zida mavjud. TRNK tuzilishi sayyoradagi barcha tirik mavjudotlarda deyarli bir xil. 2 o'lchovli ko'rinishda ko'rilganda u bargga o'xshaydi.
Biroq, agar siz uch o'lchamli ko'rinishga ega bo'lsangiz, molekula L shaklidagi geometrik tuzilishga o'xshaydi. Bu tRNKning uchinchi darajali tuzilishi hisoblanadi. Ammo o'rganish qulayligi uchun uni vizual ravishda "bo'shatish" odatiy holdir. Uchinchi darajali tuzilma ikkilamchi tuzilish elementlarining, bir-birini to'ldiruvchi qismlarning o'zaro ta'siri natijasida hosil bo'ladi.
TRNA qo'llari yoki halqalari muhim rol o'ynaydi. Bir qo'l, masalan, ma'lum bir ferment bilan kimyoviy bog'lanish uchun talab qilinadi.
Nukleotidning o'ziga xos xususiyati juda ko'p miqdordagi nukleozidlarning mavjudligi. Bu kichik nukleozidlarning 60 dan ortiq turlari mavjud.
TRNK tuzilishi va aminokislotalarni kodlash
Biz bilamizki, tRNK antikodon uzunligi 3 molekuladan iborat. Har bir antikodon o'ziga xos, "shaxsiy" aminokislotalarga mos keladi. Bu aminokislota maxsus ferment yordamida tRNK molekulasiga ulanadi. 2 ta aminokislota birikishi bilan tRNK bilan aloqalar uziladi. Barcha kimyoviy birikmalar va fermentlar talab qilinadigan vaqtdan oldin kerak bo'ladi. tRNK ning tuzilishi va funktsiyalari shu tarzda o'zaro bog'langan.
Hammasi bo'lib hujayrada bunday molekulalarning 61 turi mavjud. 64 ta matematik o'zgarishlar bo'lishi mumkin, ammo mRNKda aynan bir xil miqdordagi to'xtash kodonlari antikodonlarga ega emasligi sababli tRNKning 3 turi mavjud emas.
mRNK va tRNKning o'zaro ta'siri
Moddaning mRNK va RRNK bilan o'zaro ta'sirini, shuningdek, tRNKning tuzilish xususiyatlarini ko'rib chiqamiz. Makromolekulaning tuzilishi va maqsadi o'zaro bog'liqdir.
mRNKning tuzilishi DNKning alohida bo'limidagi ma'lumotlarni ko'chiradi. DNKning o'zi juda katta molekula birikmasidir va u hech qachon yadrodan chiqmaydi. Shuning uchun bizga vositachi RNK - axborot RNK kerak.

mRNK nusxa olgan molekulalar ketma-ketligiga asoslanib, ribosoma oqsil hosil qiladi. Ribosoma alohida polinukleotid tuzilmasi bo'lib, uning tuzilishi aniqlanishi kerak.
Ribosomal tRNK: o'zaro ta'sir
Ribosomal RNK katta organelladir. Uning molekulyar og'irligi 1 000 000 - 1 500 000 RNK umumiy miqdorining deyarli 80% ni ribosoma nukleotidlari tashkil qiladi.

Bu mRNK zanjirini ushlab, tRNK molekulalarini olib keladigan antikodonlarni kutayotganga o'xshaydi. Ribosomal RNK 2 ta subbirlikdan iborat: kichik va katta.
Ribosoma "zavod" deb ataladi, chunki kundalik hayot uchun zarur bo'lgan moddalarning barcha sintezi ushbu organellada sodir bo'ladi. Bu ham juda qadimiy hujayra tuzilishi.
Ribosomada oqsil sintezi qanday sodir bo'ladi?
tRNKning tuzilishi va uning oqsil sintezidagi roli o'zaro bog'liqdir. Ribonuklein kislotaning bir tomonida joylashgan antikodon shakli o'zining asosiy vazifasi - aminokislotalarni ribosomaga etkazib berish uchun mos keladi, bu erda oqsilning bosqichma-bosqich qurilishi sodir bo'ladi. Aslida, tRNK vositachi sifatida ishlaydi. Uning vazifasi faqat kerakli aminokislotalarni olib kelishdir.
mRNKning bir qismidan ma'lumot o'qilganda, ribosoma zanjir bo'ylab uzoqroq harakatlanadi. Matritsa faqat individual oqsilning konfiguratsiyasi va funktsiyasi haqida kodlangan ma'lumotlarni etkazish uchun kerak. Keyinchalik, azotli asoslari bo'lgan boshqa tRNK ribosomaga yaqinlashadi. Shuningdek, u mRNKning keyingi qismini dekodlaydi.
Dekodlash quyidagicha sodir bo'ladi. Azotli asoslar DNKning o'zida bo'lgani kabi komplementarlik printsipiga ko'ra birlashtiriladi. Shunga ko'ra, TRNA aminokislotalarni qayerga "bog'lash" kerakligini va qaysi "angar" ga yuborish kerakligini ko'radi.

Keyin ribosomada shu tarzda tanlangan aminokislotalar kimyoviy bog'lanadi, bosqichma-bosqich yangi chiziqli makromolekula hosil bo'ladi, u sintez tugagandan so'ng globulaga (to'pga) aylanadi. Ishlatilgan tRNK va mRNK o'z funktsiyalarini bajarib, oqsil "zavod" dan chiqariladi.
Kodonning birinchi qismi antikodon bilan juftlashganda, o'qish ramkasi aniqlanadi. Keyinchalik, agar biron sababga ko'ra ramka siljishi sodir bo'lsa, u holda ba'zi protein xususiyati rad etiladi. Ribosoma bu jarayonga aralasha olmaydi va muammoni hal qila olmaydi. Jarayon tugagandan keyingina 2 ta RRNK bo'linmasi yana birlashadi. O'rtacha har 10 4 aminokislota uchun 1 ta xato mavjud. Allaqachon yig'ilgan har 25 ta oqsil uchun kamida 1 replikatsiya xatosi bo'lishi kerak.
TRNKlar relikt molekulalar sifatida
tRNK er yuzida hayot paydo bo'lgan paytda mavjud bo'lganligi sababli, u relikt molekulasi deb ataladi. RNK DNKdan oldin mavjud bo'lgan va keyin evolyutsiyalangan birinchi tuzilma ekanligiga ishoniladi. RNK dunyosi gipotezasi - 1986 yilda laureat Valter Gilbert tomonidan ishlab chiqilgan. Biroq, buni isbotlash hali ham qiyin. Nazariya aniq faktlar bilan qo'llab-quvvatlanadi - tRNK molekulalari ma'lumotlar bloklarini saqlashga va qandaydir tarzda bu ma'lumotni amalga oshirishga, ya'ni ishlarni bajarishga qodir.
Ammo nazariya muxoliflarining ta'kidlashicha, moddaning qisqa umri tRNK har qanday biologik ma'lumotni yaxshi tashuvchisi ekanligini kafolatlay olmaydi. Bu nukleotidlar tez parchalanadi. Inson hujayralarida tRNKning umri bir necha daqiqadan bir necha soatgacha davom etadi. Ba'zi turlari bir kungacha davom etishi mumkin. Va agar bakteriyalardagi bir xil nukleotidlar haqida gapiradigan bo'lsak, unda vaqt oralig'i ancha qisqaroq - bir necha soatgacha. Bundan tashqari, tRNKning tuzilishi va funktsiyalari molekula Yer biosferasining asosiy elementiga aylanishi uchun juda murakkab.
Transport (eruvchan) RNK Translatsiya jarayonida aminokislotalarni o'sayotgan polipeptid zanjirlariga maxsus o'tkazish uchun adapter funktsiyalarini bajaradigan past molekulyar og'irlikdagi RNK molekulasi; tRNKlar shaklidagi xarakterli ikkilamchi tuzilishga ega... ...
TRNA. Eriydigan RNKga qarang. (Manba: "Genetik atamalarning inglizcha-ruscha izohli lug'ati." Arefiev V.A., Lisovenko L.A., Moskva: VNIRO nashriyoti, 1995 yil) ...
tRNK- ribonuklein kislotalarni tashish... Qisqartmalar va qisqartmalar lug'ati
Transfer RNKning tuzilishi Transfer RNK, tRNK ribonuklein kislota bo'lib, uning vazifasi aminokislotalarni sintez joyiga tashishdan iborat ... Vikipediya
Katta tibbiy lug'at
Qarang: ribonuklein kislotani tashish... Tibbiy ensiklopediya
tRNK nukleotidil transferaza- II turdagi tRNKning 3 uchiga CCA tripletini biriktiruvchi ferment (ya'ni, prekursorlarida bu triplet bo'lmagan tRNKlar, prokariotlarning ba'zi tRNKlari va aftidan, eukariotlarning barcha tRNKlari). [Arefyev V.A., Lisovenko L.A. Inglizcha-ruscha izohli lug'at...... Texnik tarjimon uchun qo'llanma
tRNKga o'xshash hudud- * tRNK o'xshash segment viruslarni o'z ichiga olgan ba'zi RNK nuklein kislotasining terminal bo'limi bo'lib, aminoatsillanish va ba'zi o'ziga xos fermentlar bilan o'zaro ta'sir qilish qobiliyatiga ega. Oddiy tRNKdan farqli o'laroq, tRNK tarkibida ... ... Genetika. ensiklopedik lug'at
tRNKga o'xshash hudud- aminokislota bilan aminokislotalar bilan aminokislotalanishga qodir va ba'zi o'ziga xos fermentlar bilan o'zaro ta'sir qilish qobiliyatiga ega bo'lgan ba'zi RNK o'z ichiga olgan viruslarning nuklein kislotasining terminal qismi; tRNKdagi tRNK dan farqli o'laroq p.u. kam uchraydigan asoslar topilmadi. ... Texnik tarjimon uchun qo'llanma
TRNK nukleotidil transferaza tRNK nukleotidil transferaza. CCA tripletini II turdagi tRNKning 3 uchiga (ya'ni, prekursorlarida bu triplet bo'lmagan tRNKlar, prokariotlarning ba'zi tRNKlari va, aftidan, eukariotlarning barcha tRNKlari) biriktiruvchi ferment.... ... Molekulyar biologiya va genetika. Izohli lug'at.
Kitoblar
- Yashirin parametrlar fizikasi: , I. Bogdanov. Ish elektr maydonlarining aylanish nazariyasi asosida yaratilgan yashirin parametrlar fizikasini tan olishga to'sqinlik qiladigan qarama-qarshiliklarni bartaraf etadi. Bor postulatlarining isboti topildi...
Darslik. tRNK ancha kichik bo'lishiga qaramay, uning tuzilishi, xususiyatlari va faoliyati haqidagi hikoya alohida bobga loyiqdir.
Shunday qilib, tRNK bir uchida genetik kodning uch harfli ketma-ketligini taniydigan "adapter" bo'lib, uni tRNKning boshqa uchiga biriktirilgan yagona mos keladigan aminokislota bilan moslashtiradi. Transfer RNKning messenjer RNKga tegishi oxirida 3 ta nukleotid hosil bo'ladi. antikodon. Faqat antikodon mRNK hududiga komplementar bo'lsa, transfer RNK unga biriktirilishi mumkin. Ammo bu holatda ham tRNK o'z-o'zidan mRNKga qo'shila olmaydi, u ularning o'zaro ta'siri joyi bo'lgan ribosomaning yordamiga, shuningdek, tarjimaning faol ishtirokchisiga muhtoj. Masalan, tRNK tomonidan olib kelingan aminokislotalar o'rtasida bog'lanish hosil qiluvchi, oqsil zanjirini hosil qiluvchi ribosomadir.
tRNKning strukturaviy xususiyatlari genetik kod bilan belgilanadi, ya'ni genga ko'ra oqsilni qurish qoidalari, RNK o'qiydi. Bu kod er yuzida yashovchi har bir jonzotda ishlaydi: virusning yaratilishi delfinning "yig'ish ko'rsatmalarini" yozish uchun ishlatiladigan bir xil uch harfli kodonlarda yozilgan. Bir tirik mavjudotning boshqasining hujayrasiga joylashtirilgan genlari mukammal tarzda ko'chirilishi va xost hujayralarida ko'payadigan genlardan farq qilmaydigan oqsillarga aylantirilishi eksperimental tarzda tasdiqlangan. Genetik kodning bir xilligi o'zgartirilgan ichak tayoqchasi koloniyalari tomonidan insulin va boshqa ko'plab inson fermentlarini ishlab chiqarish uchun asos bo'lib, ular organizmlari ularni ishlab chiqarishga qodir bo'lmagan yoki ularni etarli darajada ishlab chiqarmaydigan odamlar uchun dori sifatida ishlatiladi. Odamlar va E. coli o'rtasidagi aniq farqlarga qaramay, inson oqsillari E. coli ko'paytirish mashinasi yordamida uning dizaynlaridan osongina yaratiladi. Turli xil jonzotlarning transfer RNKlari juda kam farq qilishi ajablanarli emas.

Ushbu ro'yxatdagi har bir kodon uchtadan tashqari kodonlarni to'xtatish, tarjima tugaganligi haqida signal berib, uzatish RNK tomonidan tan olinishi kerak. Tanib olish messenjer RNKga antikodonni biriktirish orqali amalga oshiriladi, u ro'yxatdagi faqat bitta kodon bilan bog'lanishi mumkin, shuning uchun tRNK faqat bitta kodonni taniy oladi. Bu shuni anglatadiki, hujayrada ushbu molekulalarning kamida 61 turi mavjud. Aslida, ularning soni yana ham ko'p, chunki ba'zi hollarda messenjer RNKni o'qish uchun faqat kerakli antikodonga ega bo'lish etarli emas: boshqa shartlarga rioya qilish kerak, unga muvofiq maxsus, o'zgartirilgan tRNK yaratiladi. .
Bir qarashda, tRNKlarning bunday xilma-xilligi tarjima jarayonini sezilarli darajada murakkablashtirishi kerak: axir, bu molekulalarning har biri ribosoma bilan almashtirilgan xabarchi RNK kodonining antikodoniga muvofiqligini tekshiradi - bu juda ko'p ma'nosiz ko'rinadi. mexanik ish, shunchalik behuda vaqt va kuch. Ammo evolyutsiya natijasida bu muammoni oldini olish uchun uyali mexanizmlar ham rivojlandi. Misol uchun, hujayradagi har bir turdagi tRNK miqdori ushbu turdagi aminokislotalarning qurilayotgan oqsillarda qanchalik tez-tez topilganligiga mos keladi. Hujayra tomonidan kamdan-kam qo'llaniladigan aminokislotalar mavjud va tez-tez ishlatiladiganlari ham bor va agar ularni olib yuradigan tRNKlar soni bir xil bo'lsa, bu oqsillarni yig'ishni sezilarli darajada murakkablashtiradi. Shuning uchun hujayrada kamdan-kam uchraydigan aminokislotalar va ularga mos keladigan tRNKlar mavjud, tez-tez uchraydiganlari esa ko'p miqdorda ishlab chiqariladi.

Bunday xilma-xil tRNK molekulalari bilan ularning barchasi juda o'xshash, shuning uchun ularning tuzilishi va funktsiyalarini ko'rib chiqayotganda, biz asosan barcha turlarga xos xususiyatlarni o'rganamiz. tRNK ning uch o'lchovli diagrammasiga qarasangiz, u atomlarning zich aralashganiga o'xshaydi. Bu murakkab molekulaning nukleotidlarning uzun zanjirini yig'ish orqali hosil bo'lishi aql bovar qilmaydigan ko'rinadi, ammo u aynan shunday hosil bo'ladi.
Siz birinchisidan boshlab, uning hosil bo'lish bosqichlarini kuzatishingiz mumkin: ushbu transfer RNK haqidagi ma'lumotni o'z ichiga olgan genga muvofiq RNK polimeraza tomonidan nukleotidlar ketma-ketligini tuzish. Bu nukleotidlarning birin-ketin paydo bo'lish tartibi va ularning soni deyiladi tRNKning birlamchi tuzilishi. Ma'lum bo'lishicha, bu RNK polimeraza tomonidan o'qiladigan genda kodlangan tRNKning asosiy tuzilishidir. Umuman olganda, birlamchi struktura bir turdagi nisbatan sodda molekulalar ketma-ketligi bo'lib, ulardan murakkabroq, burmalangan polimer molekulasi hosil bo'ladi. Masalan, oqsil molekulasining birlamchi tuzilishi uning tarkibidagi aminokislotalarning oddiy ketma-ketligidir.

Nukleotidlarning har qanday zanjiri hujayrada ochilgan holatda bo'lolmaydi, oddiygina chiziqqa cho'zilgan. Nukleotidlarning chetlarida juda ko'p musbat va manfiy zaryadlangan qismlar mavjud bo'lib, ular bir-biri bilan vodorod aloqalarini osongina hosil qiladi. Ikki DNK molekulasining nukleotidlari o'rtasida qanday qilib bir xil aloqalar hosil bo'lib, ularni qo'sh spiralga bog'lashi tasvirlangan va vodorod aloqalari haqida batafsil ma'lumot olish uchun siz borishingiz mumkin. Vodorod bog'lari molekulalardagi atomlar orasidagi bog'lardan kamroq kuchli, ammo ular tRNK zanjirini murakkab tarzda burish va uni shu holatda ushlab turish uchun etarli. Dastlab, bu aloqalar faqat ba'zi nukleotidlar o'rtasida hosil bo'lib, tRNKni yonca bargiga o'xshash shaklga aylantiradi. Ushbu dastlabki katlamaning natijasi deyiladi ikkilamchi tuzilma tRNK. Chapdagi diagramma shuni ko'rsatadiki, faqat ba'zi nukleotidlar vodorod aloqalari bilan bog'langan, boshqalari esa juftlashtirilmagan holda halqa va halqalarni hosil qiladi. Turli xil tRNK turlarining ikkilamchi tuzilishi o'rtasidagi farq ularning birlamchi tuzilishidagi farqlarga bog'liq. Bu nukleotidlarning boshlang'ich zanjirining turli uzunliklari tufayli "yonca barglari" yoki "poyalari" ning turli uzunliklarida o'zini namoyon qiladi.
Turli tRNKlarning birlamchi tuzilishidagi yana bir farq shundaki, ular faqat ba'zi pozitsiyalarda bir xil nukleotidlarga ega (yuqoridagi diagrammada ular nomlarining birinchi harflari bilan belgilangan), lekin turli tRNKlardagi nukleotidlarning aksariyati bir-biridan farq qiladi. Yuqoridagi diagramma barcha tRNKlar uchun umumiydir, shuning uchun turli nukleotidlar raqamlar bilan belgilanadi.
tRNKning asosiy funktsional qismlari quyidagilardir:
=) antikodon, ya'ni messenjer RNKning yagona kodonini to'ldiruvchi nukleotidlar ketma-ketligi. antikodon soch turmagi
=) qabul qiluvchining oxiri, unga aminokislota biriktirilishi mumkin. U antikodon soch tolasining qarama-qarshi tomonida joylashgan.
Haqiqatda, birorta ham tRNK ikkilamchi tuzilish diagrammasidagi kabi ko'rinmaydi, chunki faqat ba'zi nukleotidlar birlashib, uni hosil qilgan, qolganlari esa juftlanmagan. Yonca bargining turli qismlaridan nukleotidlar o'rtasida vodorod aloqalari hosil bo'lishi tufayli u yanada murakkabroq bo'ladi. uchinchi darajali tuzilish L harfi shaklida. Ikkilamchi strukturaning turli qismlari qanday qilib egilib, uchinchi darajali tuzilmani hosil qilganini quyidagi diagrammalardagi ranglarni moslashtirish orqali aniq tushunishingiz mumkin. Ko'k va kulrang rangda ko'rsatilgan antikodon soch turmagi pastki qismida qoladi (esda tutish kerakki, bu "pastga" o'zboshimchalik bilan: oqsillarni tarjima qilish diagrammalarida tRNKni ushbu fazoviy yo'nalishda tasvirlash qulay) va qabul qiluvchi uchi (sariq) yon tomonga egilgan.

tRNK aminokislota biriktirishga tayyor bo'lganda shunday ko'rinadi. tRNK o'z-o'zidan aminokislota bilan birlasha olmaydi, bu maxsus fermentning ishtirokini talab qiladi: aminoatsil-tRNK sintetazalari. Hujayradagi sintetazalarning turlari soni tRNK turlarining soniga to'g'ri keladi.
Barcha turdagi tRNKlar shaklining bir xilligi ribosoma ularning har qandayini tanib olishi, mRNK bilan bog'lanishini osonlashtirishi va ularni bir joydan ikkinchisiga o'tkazishi uchun zarurdir. Agar turli xil tRNK turlari bir-biridan sezilarli darajada farq qilsa, bu ribosoma ishini juda qiyinlashtirib, oqsil sintezi tezligini keskin kamaytiradi. Shunday qilib, tabiiy tanlanish tRNKlarni bir-biriga o'xshash qilishni maqsad qiladi. Ammo shu bilan birga, tRNKning har xil turlari o'rtasida sezilarli farqlar mavjudligini talab qiladigan yana bir omil mavjud: axir, har bir turni tanib olish va unga mos keladigan yagona aminokislotalarni biriktirish kerak. Shubhasiz, bu farqlar sezilarli bo'lishi kerak, lekin juda muhim emas, shuning uchun tRNK turlarini tanib olish ishi zargarlik jarayoniga aylanadi. Aminoatsil-tRNK sintetazlari aynan shunday qiladi: ularning har biri 20 ta aminokislotadan faqat bittasi bilan aloqa qilishi va uni aynan shu aminokislotaga mos keladigan tRNK turlariga biriktirishi mumkin. Genetik kod ko'rsatilgan jadvaldan ko'rinib turibdiki, har bir aminokislota bir nechta nukleotidlar ketma-ketligi bilan kodlangan, shuning uchun, masalan, CGA, CGG, CGU va CGC antikodonlari bo'lgan to'rtta tRNKning barchasi alaninni biriktiradigan bir xil sintetaza tomonidan tan olinadi. ular. Bitta sintetaza tomonidan qayta ishlangan bunday tRNKlar deyiladi bog'liq.
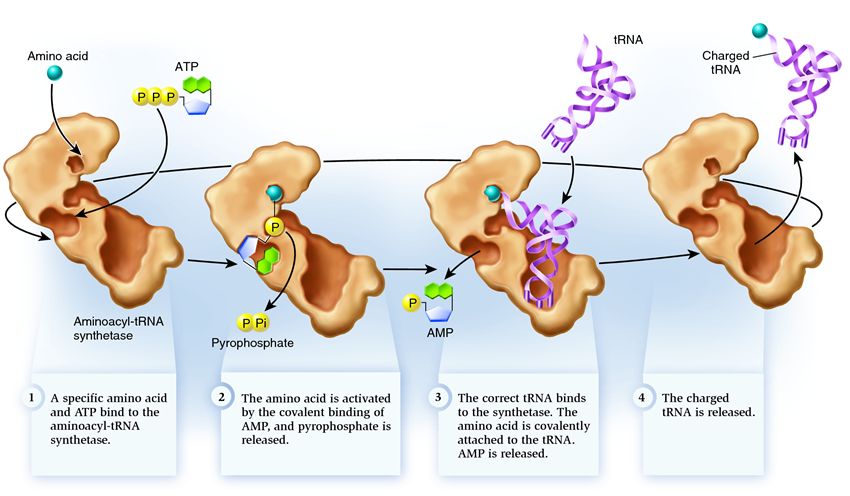
Sintetaza fermentlar guruhiga kiradi, ularning vazifasi alohida mavjud molekulalar bilan bog'lanish va ularni bittaga birlashtirish:
1 . sintetaza aminokislota va ATP molekulasini bog'laydi. Ikki fosfat guruhi ATP dan ajralib, keyingi qadamlar uchun zarur bo'lgan energiyani chiqaradi. Vayron bo'lgan molekuladan qolgan adenozin monofosfat (AMP) aminokislotalarga biriktirilib, uni qabul qiluvchi soch ipi bilan bog'lanishga tayyorlaydi.
2 . sintetaza o'ziga shu aminokislotaga mos keladigan tegishli tRNKlardan birini biriktiradi.

Ushbu bosqichda transfer RNK sintetazasining muvofiqligi tekshiriladi. Bir nechta tanib olish usullari mavjud va har bir sintetaza ularning o'ziga xos kombinatsiyasiga ega. Antikodonning kamida bitta nukleotidi sintetaza va tRNKning o'zaro ta'sirida ishtirok etadi. Akseptor soch ipi ham tekshirishni talab qiladi: unda kerakli aminokislotaga mos keladigan tegishli tRNKlar uchun umumiy bo'lgan o'ziga xos nukleotidlar mavjudligi aniqlanadi. tRNKning boshqa qismlarining nukleotidlari ham sintetazaning ma'lum joylariga bog'lanib, moslashishda ishtirok etishi mumkin. Noto'g'ri tRNK ba'zi parametrlarda kerakli bilan mos kelishi mumkin, ammo to'liq bo'lmaganligi sababli u sintetazaga sekin va bo'shashmasdan biriktiriladi, osongina tushib ketadi. Va to'g'ri tRNK sintetazaga tez va mahkam yopishadi, buning natijasida sintetaza tuzilishi o'zgaradi va jarayon boshlanadi. aminoatsillanish , ya'ni tRNKga aminokislota qo'shilishi.

3 . Aminoatsillanish aminokislotaga biriktirilgan AMP molekulasini tRNK molekulasi bilan almashtirishni o'z ichiga oladi. Ushbu almashtirilgandan so'ng, AMP sintetazadan chiqib ketadi va tRNK aminokislotalarni yakuniy tekshirish uchun ushlab turiladi. Agar biriktirilgan aminokislota noto'g'ri ekanligi aniqlansa, u tRNKdan ajralib chiqadi va aminokislotalarning sintetazadagi o'rni bo'sh qoladi va u erga boshqa molekula qo'shilishi mumkin. Yangi aminokislota ATP va tRNK bilan birlashish bosqichlaridan o'tadi va sinovdan o'tadi. Agar xatolikka yo'l qo'yilmasa, aminokislota bilan zaryadlangan tRNK chiqariladi: u oqsillarni tarjima qilishda o'z rolini o'ynashga tayyor. Va sintetaza yangi aminokislotalar va tRNKlarni biriktirishga tayyor va tsikl yana boshlanadi.
Ko'p narsa aminoatsil-tRNK sintetazasining to'g'ri ishlashiga bog'liq: agar bu bosqichda nosozlik yuzaga kelsa, tRNKga noto'g'ri aminokislotalar biriktiriladi. Va u ribosomada o'sadigan oqsilga o'rnatiladi, chunki tRNK va ribosoma kodon va aminokislotalarning mosligini tekshirish funktsiyasiga ega emas. Xatoning oqibatlari kichik yoki halokatli bo'lishi mumkin va tabiiy tanlanish jarayonida bunday tekshiruvlar funktsiyasiga ega bo'lmagan fermentlarga ega mavjudotlar aminokislotalar va tRNK o'rtasidagi yozishmalarni o'rnatish uchun turli xil variantlarga ega bo'lgan ko'proq moslashuvchan mavjudotlar bilan almashtirildi. . Shuning uchun zamonaviy hujayralarda sintetaza noto'g'ri aminokislota bilan o'rtacha 50 mingtadan bittasida, noto'g'ri tRNK bilan esa 100 ming ulanishda bir marta birlashadi.

Ba'zi aminokislotalar bir-biridan faqat bir nechta atomlar bilan farq qiladi. Agar siz ularning diagrammalariga qarasangiz, argininni alanin bilan chalkashtirib yuborish ehtimoli izolösinni leysin yoki valin bilan adashtirishdan ko'ra kamroq ekanligi ayon bo'ladi. Shuning uchun bir-biriga o'xshash aminokislotalardan biriga bog'langan har bir sintetaza qo'shimcha tekshirish mexanizmlariga ega. Mana izolösin-tRNK sintetazadagi bunday moslashuvga misol:

Har bir sintetaza mavjud sintetik markaz, unda tRNKga aminokislota qo'shiladi. Sintetaza tomonidan ushlangan tRNKning akseptor soch tolasi u bilan reaksiyaga kirishishga tayyor bo'lgan aminokislota kabi tugaydi. Ba'zi sintetazalarning ishi aminokislota va tRNK birlashgandan so'ng darhol tugaydi. Ammo Ile-tRNK sintetaza izolösinga o'xshash boshqa aminokislotalar mavjudligi sababli xato qilish ehtimolini oshiradi. Shuning uchun u ham bor axloq tuzatish markazi: Nomidan tRNK va aminokislotalarni bog'lash jarayonida qanday rol o'ynashi aniq. O'ngdagi rasmda akseptor tRNK soch tolasi uchining Ile-tRNK sintetazasining sintetik markazida joylashishi bu soch tolasiga g'ayritabiiy egilish hosil qilishini ko'rsatadi. Biroq, sintetaza tRNK ni shu holatda unga aminokislota qo'shilmaguncha ushlab turadi. Ushbu bog'lanish sodir bo'lgandan so'ng, sintetik markazda akseptor soch turmagiga bo'lgan ehtiyoj tugaydi va tRNK to'g'rilanadi va oxiri tuzatish markazida unga biriktirilgan aminokislota bilan tugaydi.

Albatta, sintetik markaz sintetaza uchun mos bo'lmagan aminokislotalarni tekshirishda ham rol o'ynaydi. Unga kirish uchun molekula bir qator shartlarga javob berishi kerak, jumladan, kerakli o'lchamga ega bo'lishi kerak. Leysin va izolösin bir xil miqdordagi atomlarni o'z ichiga olgan bo'lsa-da, fazoviy tuzilishdagi farqlar tufayli leytsin kattaroqdir. Shuning uchun u o'lchami ixchamroq izolösinga to'g'ri keladigan sintetik markazga kira olmaydi va shunchaki Ile-tRNK sintetazasidan sakrab chiqadi.
Ammo atom tuzilishi oʻxshash boʻlgan bu uch molekula ichida eng kichigi boʻlgan valin sintetik markazda izolösin oʻrnini osongina egallaydi va sintetaza uni tRNKga biriktiradi. Bu erda sintetaza tuzatish markazi o'ynaydi. Agar to'g'rilash qabul qiluvchi soch tolasi to'g'ri zaryadlangan bo'lsa va izolösinni olib yursa, u tuzatish markazining ichida siqib chiqa olmaydi: bu molekula uchun juda kichik. Shunday qilib, endi hech narsa to'g'rilangan tRNKni ushlab turmaydi va u sintetazadan ajratiladi. Ammo agar valin tRNKga biriktirilgan bo'lsa, u korreksiya markaziga o'tadi va shu bilan u bilan bog'langan tRNKni sintetazada ushlab turadi. Ichkarida tRNKning bunday haddan tashqari uzoq bo'lishi sintetaza uchun xato signali bo'lib, uning fazoviy konfiguratsiyasini o'zgartiradi. Natijada:
=) valin tRNK dan ajratiladi va sintetazadan chiqariladi
=) akseptor soch tolasi aminokislota bilan biriktirilishini kutib, sintetik markazga qaytadi
=) sintetaza yangi aminokislota bilan bog'lanadi, u bilan tRNKni "zaryadlaydi" va buning uchun izolösin ishlatilgan yoki yo'qligini yana tekshiradi.
Shunga o'xshash ikki tomonlama tanib olish mexanizmi boshqa sintetazlar tomonidan qo'llaniladi.
Aminoatsil-tRNK sintetaza (ARCase) sintetaza fermenti bo'lib, ma'lum bir aminokislotaning tegishli tRNK molekulasi bilan esterifikatsiya reaktsiyasida aminoatsil-tRNK hosil bo'lishini katalizlaydi. Har bir aminokislota o'zining aminoatsil-tRNK sintetazasiga ega. ARSazalar oqsil tarkibiga kiritilgan aminokislotalarning genetik kodining (tRNK antikodon) nukleotid tripletlari bilan mos kelishini ta'minlaydi va shu bilan ribosomalarda oqsil sintezi jarayonida mRNKdan genetik ma'lumotni keyingi o'qishning to'g'riligini ta'minlaydi. Aksariyat APCazalar 1, 2 yoki 4 ta bir xil polipeptid zanjirlaridan iborat. Polipeptid zanjirlarining molekulyar og'irligi 30-140 ming. Ko'p APCazlar ikkita faol markazni o'z ichiga oladi. 3 ta uchastka mavjud. Birinchi mintaqaning o'ziga xos xususiyati yo'q, u barcha fermentlar uchun bir xildir; Ikkinchi mintaqa qat'iy o'ziga xoslikka ega, bu erda ma'lum bir AK biriktiriladi, shuning uchun u ARSaz deb ataladi, masalan, metioninni biriktirsa, u metionil-tRNK sintetaza deb ataladi; Uchinchi mintaqa ham qat'iy o'ziga xos mintaqadir va faqat ma'lum bir tRNK bilan bog'lanishi mumkin. Shunday qilib, ferment aminokislotalar va tRNKni tanib olish uchun zarurdir.
APCaselar tomonidan katalizlanadigan reaktsiyalarning o'ziga xosligi juda yuqori bo'lib, bu tirik hujayradagi oqsil sintezining aniqligini belgilaydi. Agar A. tuzilishi oʻxshash aminokislotaga ega tRNKning notoʻgʻri aminoatsillanishini amalga oshirsa, bir xil APCase tomonidan katalizlangan notoʻgʻri AA-tRNKlarning AA va tRNKga gidrolizlanishi orqali korreksiya sodir boʻladi. Sitoplazmada xloroplastlarning to'liq to'plami mavjud va mitoxondriyalar o'zlarining APCazlariga ega;
RNKni uzatish. Tuzilishi, funktsiyalari. Ribosomalarning tuzilishi.
Barcha tRNKlar birlamchi tuzilishida ham, nukleotid qoldiqlari asoslari oʻrtasidagi oʻzaro taʼsir natijasida polinukleotid zanjirining ikkilamchi tuzilishga burmalanishida ham umumiy xususiyatlarga ega.
tRNKning birlamchi tuzilishi
tRNKlar nisbatan kichik molekulalar bo'lib, ularning zanjirlarining uzunligi 74 dan 95 gacha nukleotid qoldiqlari orasida o'zgarib turadi. Barcha tRNKlar ikkita sitozin qoldig'i va bitta adenozin qoldig'idan (CCA end) qurilgan bir xil 3" uchiga ega. Bu aminokislota qoldig'i bilan aminoatsil-tRNK hosil bo'lishida bog'langan 3" terminal adenozindir. CCA uchi ko'plab tRNKlarga maxsus ferment yordamida biriktirilgan. Aminokislota (antikodon) uchun kodonni to'ldiruvchi nukleotid tripleti taxminan tRNK zanjirining o'rtasida joylashgan. Ketma-ketlikning ma'lum pozitsiyalarida deyarli barcha turdagi tRNKlar bir xil (saqlangan) nukleotid qoldiqlarini o'z ichiga oladi. Ba'zi pozitsiyalarda faqat purin yoki faqat pirimidin asoslari bo'lishi mumkin (ular yarim konservativ qoldiqlar deb ataladi).
Barcha tRNK molekulalari ko'p miqdorda (barcha qoldiqlarning 25% gacha) turli xil modifikatsiyalangan nukleozidlarning mavjudligi bilan tavsiflanadi, ko'pincha kichik deb ataladi. Ular molekulalarning turli joylarida, ko'p hollarda aniq belgilangan, oddiy nukleozid qoldiqlarini maxsus fermentlar yordamida o'zgartirish natijasida hosil bo'ladi.
tRNKning ikkilamchi tuzilishi
zanjirning ikkilamchi tuzilishga katlanishi zanjir bo'limlarining o'zaro to'ldirilishi tufayli sodir bo'ladi. Uchta zanjir bo'lagi o'z-o'zidan buklanganda bir-birini to'ldiruvchi bo'lib, soch turmagiga o'xshash tuzilmalarni hosil qiladi. Bundan tashqari, 5" uchi zanjirning 3" uchiga yaqin bo'lgan hududni to'ldiradi, ularning antiparallel joylashuvi bilan; ular qabul qiluvchi poya deb ataladigan poyani hosil qiladi. Natijada, "yonda bargi" deb ataladigan to'rtta poya va uchta halqa mavjudligi bilan tavsiflangan tuzilish paydo bo'ladi. Poya va halqa shoxcha hosil qiladi. Pastki qismida antikodon shoxchasi joylashgan bo'lib, uning halqasining bir qismi sifatida antikodon tripleti mavjud. Buning chap va o'ng tomonida D va T shoxlari joylashgan bo'lib, ularning halqalarida g'ayrioddiy saqlanib qolgan nukleozidlar dihidroridin (D) va timidin (T) mavjudligi sababli nomlangan. O'rganilgan barcha tRNKlarning nukleotidlar ketma-ketligi o'xshash tuzilmalarga katlanishi mumkin. Uchta yonca bargi halqasidan tashqari tRNKda qoʻshimcha yoki oʻzgaruvchan halqa (V-loop) ham mavjud. Uning o'lchamlari turli xil tRNKlar orasida keskin farq qiladi, 4 dan 21 nukleotidgacha va oxirgi ma'lumotlarga ko'ra, 24 nukleotidgacha.
tRNKning fazoviy (uchlamchi) tuzilishi
Ikkilamchi tuzilish elementlarining o'zaro ta'siri tufayli uchinchi darajali tuzilma hosil bo'ladi, u lotincha L harfiga o'xshashligi sababli L-shakl deb ataladi (2 va 3-rasm). Asosiy stacking orqali qabul qiluvchi poya va yonca bargi T poyasi bitta uzluksiz qo‘sh spirali, qolgan ikkita poya antikodon va D esa yana bir uzluksiz qo‘sh spirali hosil qiladi. Bunday holda, D- va T-looplar bir-biriga yaqinlashadi va qo'shimcha, ko'pincha g'ayrioddiy, tayanch juftlarini shakllantirish orqali bir-biriga mahkamlanadi. Konservativ yoki yarim konservativ qoldiqlar, qoida tariqasida, bu juftlarning shakllanishida ishtirok etadi. Shunga o'xshash uchinchi darajali o'zaro ta'sirlar L-tuzilmasining boshqa qismlarini birlashtiradi
Transfer RNK (tRNK) ning asosiy maqsadi ribosomaga faollashtirilgan aminokislota qoldiqlarini yetkazib berish va matritsadagi genetik kod yoki ma'lumot, RNK (mRNK) tomonidan yozilgan dasturga muvofiq sintezlangan oqsil zanjiriga kiritilishini ta'minlashdir.
Ribosomalarning tuzilishi.
Ribosomalar - bu ribonukleoprotein hosilalari - aminokislotalar oqsillarga birikadigan "zavod" turi. Eukaryotik ribosomalar cho‘kma konstantasi 80S bo‘lib, 40S (kichik) va 60S (katta) bo‘linmalardan iborat. Har bir subbirlik rRNK va oqsillarni o'z ichiga oladi.
Proteinlar bir nusxada ribosoma bo'linmalarining bir qismi bo'lib, aminokislota yoki peptid bilan bog'langan mRNK va tRNK o'rtasidagi o'zaro ta'sirni ta'minlovchi tizimli funktsiyani bajaradi.
mRNK ishtirokida 40S va 60S subbirliklari birlashib, toʻliq ribosomani hosil qiladi, uning ogʻirligi gemoglobin molekulasining massasidan taxminan 650 baravar katta.
Ko'rinib turibdiki, rRNK ribosomalarning asosiy strukturaviy va funktsional xususiyatlarini aniqlaydi, xususan, ribosoma bo'linmalarining yaxlitligini ta'minlaydi, ularning shakli va bir qator tuzilish xususiyatlarini aniqlaydi.
Katta va kichik bo'linmalarning birlashishi xabarchi RNK (mRNK) ishtirokida sodir bo'ladi. Bir mRNK molekulasi odatda bir nechta ribosomalarni munchoqlar qatori kabi bir-biriga bog'laydi. Ushbu tuzilish polisoma deb ataladi. Polisomalar sitoplazmaning asosiy moddasida erkin joylashadi yoki qo'pol sitoplazmatik retikulumning membranalariga biriktiriladi. Ikkala holatda ham ular faol oqsil sintezi joyi bo'lib xizmat qiladi.
Endoplazmatik retikulum singari ribosomalar ham faqat elektron mikroskop yordamida topilgan. Ribosomalar hujayra organellalarining eng kichik qismidir.
Ribosomada tRNK molekulalarining biriktirilishi uchun 2 ta markaz mavjud: aminoatsil (A) va peptidil (P) markazlari, ularning shakllanishida ikkala subbirlik ham ishtirok etadi. A va P markazlari birgalikda 2 ta kodonga teng mRNK hududini o'z ichiga oladi. Tarjima paytida A markazi aa-tRNKni bog'laydi, uning tuzilishi ushbu markaz hududida joylashgan kodon tomonidan belgilanadi. Ushbu kodonning tuzilishi o'sib borayotgan polipeptid zanjiriga qo'shiladigan aminokislotalarning tabiatini kodlaydi. P markazini peptidil-tRNK egallaydi, ya'ni. tRNK allaqachon sintez qilingan peptid zanjiri bilan bog'langan.
Eukariotlarda ikki xil ribosomalar mavjud: hujayralar sitoplazmasida joylashgan "erkin" va endoplazmatik retikulum (ER) bilan bog'langan. ER bilan bog'liq ribosomalar qon plazmasiga chiqariladigan va ER oqsillari, Golji apparati membranasi, mitoxondriyalar yoki lizosomalarning yangilanishida ishtirok etadigan "eksport uchun" oqsillarni sintez qilish uchun javobgardir.
Polipeptid molekulasining sintezi. Boshlanish va cho'zilish.
Protein sintezi tsiklik, ko'p bosqichli, energiyaga bog'liq jarayon bo'lib, unda erkin aminokislotalar polipeptidlarni hosil qilish uchun genetik jihatdan aniqlangan ketma-ketlikda polimerlanadi.
Matritsa oqsili sintezining ikkinchi bosqichi, ribosomada sodir bo'ladigan haqiqiy tarjima shartli ravishda uch bosqichga bo'linadi: boshlash, cho'zilish va tugatish.
Boshlash.
Yagona mRNKga transkripsiyalangan DNK ketma-ketligi 5' uchida qidirishdan boshlanib, 3' uchida terminator bilan tugaydi, transkripsiya birligi bo'lib, "gen" tushunchasiga mos keladi. Gen ifodasini nazorat qilish tarjimani boshlash bosqichida amalga oshirilishi mumkin. Ushbu bosqichda RNK polimeraza promotorni - 41-44 bp uzunlikdagi fragmentni taniydi. Transkripsiya 5`-3` yo'nalishda yoki chapdan o'ngga sodir bo'ladi. tRNK sintezi boshlanadigan boshlang'ich nukleotidning o'ng tomonida joylashgan ketma-ketliklar + belgisi bilan (+1,+2..) va chapda - (-1,-2) belgisi bilan raqamlar bilan belgilanadi. Shunday qilib, DNK polimeraza biriktirilgan DNK hududi koordinatalari taxminan -20 dan +20 gacha bo'lgan hududni egallaydi. Barcha promotorlar bir xil nukleotidlar ketma-ketligini o'z ichiga oladi, ular saqlanib qolgan deb ataladi. Bunday ketma-ketliklar RNK polimerazalari tomonidan tan olingan signallar bo'lib xizmat qiladi. Boshlanish nuqtasi odatda purin hisoblanadi. Buning darhol chap tomonida 6-9 bp Pribnow ketma-ketligi (yoki quti) sifatida tanilgan: TATAAT. U biroz farq qilishi mumkin, lekin birinchi ikkita asos ko'pchilik promouterlarga kiritilgan. Taxminlarga ko'ra, u ikkita vodorod aloqasi bilan bog'langan AT juftlariga boy mintaqa tomonidan yaratilganligi sababli, bu joydagi DNK alohida zanjirlarga osonroq bo'linadi. Bu RNK polimeraza ishlashi uchun sharoit yaratadi. Shu bilan birga, Pribnow qutisi mRNK sintezi chapdan o'ngga, ya'ni 5`-3` gacha davom etadigan tarzda yo'naltirish uchun zarurdir. Pribnov qutisining markazi nukleotid -10 da joylashgan. Shunga o'xshash tarkibdagi ketma-ketlik 35-pozitsiyada markazlashtirilgan boshqa mintaqada joylashgan. 9 bp dan iborat ushbu mintaqa 35-ketma yoki tanib olish hududi deb belgilangan. Bu omil bog'langan joy bo'lib, shu bilan RNK polimeraza maxsus oqsillarsiz transkripsiyani boshlay olmaydigan samaradorlikni aniqlaydi. Ulardan biri CAP yoki CRP omilidir.
Eukariotlarda RNK polimeraza II bilan o'zaro ta'sir qiluvchi promotorlar batafsilroq o'rganilgan. Ular koordinatalari -25, -27 nuqtalarda, shuningdek, boshlang'ich nuqtalarda joylashgan uchta gomologik bo'limni o'z ichiga oladi. Boshlangʻich asoslari adenin boʻlib, uning ikki tomonida pirimidinlar joylashgan. 19-25 bp masofada. 7 bp saytning chap tomonida joylashgan. TATA ketma-ketligi yoki Hogness qutisi sifatida tanilgan TATAA ko'pincha GC juftlariga boy hududlar bilan o'ralgan. Hatto chap tomonda -70 dan -80 gacha bo'lgan pozitsiyalarda CAAT qutisi deb ataladigan GTZ yoki CAATCT ketma-ketligi joylashgan. Taxminlarga ko'ra, TATA ketma-ketligi boshlang'ich nukleotidni tanlashni, CAAT esa RNK polimerazasining DNK shabloniga birlamchi ulanishini nazorat qiladi.
Cho'zilish. mRNK cho'zilish bosqichi DNKning cho'zilishiga o'xshaydi. U prekursorlar sifatida ribonukleotid trifosfatlarni talab qiladi. Transkripsiya uzayish bosqichi, ya'ni mRNK zanjirining o'sishi pirofosfat ajralib chiqishi bilan zanjirning 3'-uchiga ribonukleotid monofosfatlarning biriktirilishi bilan sodir bo'ladi. Eukariotlarda nusxa ko'chirish odatda DNKning (genning) cheklangan qismida sodir bo'ladi, ammo prokariotlarda ba'zi hollarda transkripsiya bitta operon va bitta umumiy promotorni tashkil etuvchi bir nechta bog'langan genlar orqali ketma-ket sodir bo'lishi mumkin. Bunday holda, polikistronik mRNK hosil bo'ladi.
Laktoza operoni misolida gen faolligini tartibga solish.
Laktoza operoni bakteriyalarning polikistronik operoni bo'lib, laktoza metabolizmi uchun genlarni kodlaydi.
Ichak tayoqchasida laktoza almashinuvi genlarining ifodalanishini tartibga solish birinchi marta 1961 yilda olimlar F. Yakob va J. Monod tomonidan tasvirlangan. Bakterial hujayra laktoza almashinuvida ishtirok etuvchi fermentlarni faqat atrof-muhitda laktoza mavjud bo'lganda va hujayrada glyukoza bo'lmasa sintez qiladi.
Laktoza operoni uchta strukturaviy gen, promotor, operator va terminatordan iborat. Taxminlarga ko'ra, operon repressor oqsilni kodlaydigan regulyator genini ham o'z ichiga oladi.
Laktoza operonining strukturaviy genlari - lacZ, lacY va lacA:
lacZ laktoza disaxaridini glyukoza va galaktozaga parchalaydigan b-galaktozidaza fermentini kodlaydi;
lacY laktozani hujayra ichiga olib o'tuvchi membrana transport oqsili b-galaktozid o'tkazuvchanligini kodlaydi.
lacA atsetil guruhini asetil-KoA dan beta-galaktozidlarga o'tkazuvchi ferment b-galaktozid transatsetilazani kodlaydi.
Har bir operonning boshida maxsus gen - operator geni mavjud. Bitta m-RNK odatda bitta operonning strukturaviy genlarida hosil bo'ladi va bu genlar bir vaqtning o'zida faol yoki harakatsiz bo'lishi mumkin. Qoida tariqasida, operondagi strukturaviy genlar repressiya holatidadir.
Promotor - DNKning RNK polimeraza fermenti tomonidan tan olingan bo'limi bo'lib, u operonda m-RNK sintezini ta'minlaydi, undan oldin DNKning faollashtiruvchi oqsili bo'lgan Sar oqsili biriktirilgan qismi joylashgan. DNKning bu ikki qismi 85 juft nukleotiddan iborat. Promotordan keyin operon 21 juft nukleotiddan iborat bo'lgan operator genini o'z ichiga oladi, odatda regulyator gen tomonidan ishlab chiqarilgan repressor oqsili operator genining orqasida bo'shliq (bo'shliq) mavjud. Spacerlar DNK molekulasining turli uzunlikdagi (ba'zan 20 000 ta asosiy juftlik) ma'lumotga ega bo'lmagan qismlari bo'lib, ular qo'shni genning transkripsiya jarayonini tartibga solishda ishtirok etadilar.
Operon terminator bilan tugaydi - bu operonda m-RNK sintezi uchun to'xtash signali bo'lib xizmat qiluvchi DNKning kichik qismi.
Qabul qiluvchi genlar strukturaviy genlarning faoliyatini tartibga soluvchi turli oqsillarni biriktiruvchi joy bo'lib xizmat qiladi. Agar laktoza hujayra ichiga kirib boradigan bo'lsa (bu holda u induktor deb ataladi) regulyator gen tomonidan kodlangan oqsillarni blokirovka qilsa, u holda ular operator geniga ulanish qobiliyatini yo'qotadi. Operator geni faol holatga o'tadi va strukturaviy genlarni ishga tushiradi.
RNK polimeraza, Cap oqsili (aktivator oqsili) yordamida promotorga birikadi va operon bo'ylab harakatlanib, pro-m-RNKni sintez qiladi. Transkripsiya paytida m-RNK bitta operondagi barcha strukturaviy genlardan genetik ma'lumotni o'qiydi. Ribosomaga tarjima qilish jarayonida bir nechta turli xil polipeptid zanjirlari m-RNK tarkibidagi kodonlarga muvofiq sintezlanadi - har bir zanjirning tarjimasining boshlanishi va tugashini ta'minlaydigan nukleotidlar ketma-ketligi. Laktoza operoni misolida ko'rib chiqilgan gen funktsiyasini tartibga solish turi oqsil sintezining salbiy induksiyasi deb ataladi.
Triptofan operoni misolida gen faolligini tartibga solish.
Genlarni tartibga solishning yana bir turi - E.coUda aminokislotalar triptofonining sintezini boshqaradigan operon misolida o'rganilgan salbiy repressiya. Ushbu operon 6700 juft nukleotiddan iborat bo'lib, 5 ta strukturaviy gen, operator geni va ikkita promotorni o'z ichiga oladi. Regulyator geni trp operonning ishlashiga ta'sir qilmaydigan tartibga soluvchi oqsilning doimiy sintezini ta'minlaydi. Hujayrada triptofan ko'p bo'lsa, ikkinchisi tartibga soluvchi oqsil bilan bog'lanadi va uni operon bilan bog'laydigan va mos keladigan m-RNK sintezini bostiradigan tarzda o'zgartiradi.
Genetik faoliyatning salbiy va ijobiy nazorati.
Ijobiy induksiya deb ataladigan narsa ham ma'lum, regulyator genining oqsil mahsuloti operonning ishlashini faollashtirganda, ya'ni. repressor emas, balki faollashtiruvchi bu bo'linish shartli bo'lib, operonning akseptor qismining tuzilishi va prokariotlarda gen regulyatorining ta'siri juda xilma-xildir.
Prokaryotlarda operondagi strukturaviy genlar soni birdan o'n ikkigacha; Operonda bir yoki ikkita promouter va terminator bo'lishi mumkin. Bitta operonda lokalizatsiya qilingan barcha strukturaviy genlar, qoida tariqasida, biokimyoviy reaktsiyalarning bir zanjirini ta'minlaydigan fermentlar tizimini boshqaradi. Hujayrada bir nechta operonlarni tartibga solishni muvofiqlashtiruvchi tizimlar mavjudligiga shubha yo'q.
m-RNK sintezini faollashtiruvchi oqsillar gen akseptorining birinchi qismiga - operatorga, uning oxiriga esa m-RNK sintezini bostiruvchi repressor oqsillarga biriktiriladi. Bitta gen bir nechta oqsillardan biri tomonidan boshqariladi, ularning har biri mos keladigan qabul qiluvchi joyga biriktiriladi. Turli xil genlar umumiy regulyatorlarga va bir xil operator hududlariga ega bo'lishi mumkin. Gen regulyatorlari bir vaqtning o'zida harakat qilmaydi. Birinchidan, biri darhol bir guruh genlarni ishga tushiradi, keyin bir muncha vaqt o'tgach, ikkinchisi boshqa guruhga aylanadi, ya'ni. gen faolligini tartibga solish "kaskadlar" da sodir bo'ladi va bir bosqichda sintez qilingan oqsil keyingi bosqichda oqsil sintezining regulyatori bo'lishi mumkin.
Xromosomalarning tuzilishi. Karyotip. Idiogramma. Xromosoma tuzilishi modellari.
Eukaryotik xromosomalar murakkab tuzilishga ega. Xromosomaning asosini sezilarli uzunlikdagi dezoksiribonuklein kislotasining (DNK) chiziqli (halqada yopilmagan) makromolekulasi tashkil etadi (masalan, inson xromosomalarining DNK molekulalarida 50 dan 245 million juft azotli asoslar mavjud). Cho'zilganida, inson xromosomasining uzunligi 5 sm ga etishi mumkin, unga qo'shimcha ravishda xromosoma beshta maxsus oqsillarni - H1, H2A, H2B, H3 va H4 (gistonlar deb ataladi) va bir qator giston bo'lmagan oqsillarni o'z ichiga oladi. . Gistonlarning aminokislotalar ketma-ketligi yuqori darajada saqlanadi va organizmlarning eng xilma-xil guruhlarida amalda farq qilmaydi. Interfazada xromatin kondensatsiyalanmaydi, lekin bu vaqtda ham uning iplari DNK va oqsillar majmuasidir. Xromatin deoksiribonukleoprotein bo'lib, yorug'lik mikroskopida ingichka iplar va granulalar shaklida ko'rinadi. DNK makromolekulasi H2A, H2B, H3 va H4 giston oqsillarining oktomerlarini (sakkizta oqsil globulasidan tashkil topgan tuzilmalar) o'rab, nukleosomalar deb ataladigan tuzilmalarni hosil qiladi.
Umuman olganda, butun tuzilish biroz boncuklarni eslatadi. H1 oqsili bilan bog'langan bunday nukleosomalar ketma-ketligi diametri taxminan 10 nm bo'lgan nukleofilament yoki nukleosoma ipi deb ataladi.
Kondensatsiyalangan xromosoma X shakliga ega (ko'pincha teng bo'lmagan qo'llar bilan), chunki replikatsiya natijasida hosil bo'lgan ikkita xromatid hali ham sentromerada bog'langan. Inson tanasining har bir hujayrasida aniq 46 ta xromosoma mavjud. Xromosomalar har doim juftlashgan. Hujayrada har doim har bir turdagi 2 ta xromosoma bo'ladi.
Tsentromera - bu har ikkala opa-singil xromatidaga xos bo'lgan xromosomaning maxsus tashkil etilgan hududi. Tsentromera xromosoma tanasini ikki qismga ajratadi. Birlamchi konstriksiyaning joylashishiga qarab, xromosomalarning quyidagi turlari ajratiladi: teng qurolli (metasentrik), sentromera o'rtada joylashganda va qo'llar uzunligi taxminan teng bo'lganda; teng bo'lmagan qo'llar (submetasentrik), sentromera xromosoma o'rtasidan siljiganida va qo'llar teng bo'lmagan uzunlikda; tayoqchali (akrotsentrik), sentromera xromosomaning bir uchiga siljiganida va bir qo'li juda qisqa bo'ladi. Ba'zi xromosomalarda yo'ldosh deb ataladigan hududni xromosoma tanasidan ajratib turadigan ikkilamchi siqilishlar bo'lishi mumkin.
Eukaryotik hujayralardagi xromosomalarning kimyoviy tuzilishini o'rganish ular asosan DNK va oqsillardan iborat ekanligini ko'rsatdi. Ko'pgina tadqiqotlar tomonidan tasdiqlanganidek, DNK irsiyat va o'zgaruvchanlik xususiyatlarining moddiy tashuvchisi bo'lib, biologik ma'lumotlarni o'z ichiga oladi - maxsus kod yordamida qayd etilgan hujayra yoki organizmni rivojlantirish dasturi. Proteinlar xromosomalar moddasining muhim qismini (bu tuzilmalar massasining taxminan 65%) tashkil qiladi. Genlar majmuasi sifatida xromosoma ma'lum bir turning barcha individlariga xos bo'lgan evolyutsion tarzda o'rnatilgan tuzilishdir. Xromosoma ichidagi genlarning nisbiy joylashuvi ularning faoliyati tabiatida muhim rol o'ynaydi.
Karyotipning strukturaviy xususiyatlarini ko'rsatuvchi grafik tasviri idiogramma deyiladi.
Xromosomalarning soni va tuzilishi bo'yicha ma'lum bir turga xos bo'lgan xromosomalar to'plamiga karyotip deyiladi.
Gistonlar. Nukleosoma tuzilishi.
Gistonlar - nukleoproteinlarning asosiy sinfi, DNK zanjirlarini xromosomalarga yig'ish va qadoqlash uchun zarur bo'lgan yadro oqsillari. H1/H5, H2A, H2B, H3, H4 nomli besh xil giston turi mavjud. Ushbu oqsillardagi aminokislotalarning ketma-ketligi turli darajadagi tashkiliy organizmlarda deyarli farq qilmaydi. Gistonlar kichik, juda asosli oqsillar bo'lib, ular bevosita DNK bilan bog'lanadi. Gistonlar aminokislotalar qoldiqlarining musbat zaryadlari hisobiga DNKning manfiy zaryadlangan fosfat guruhlarini zararsizlantirib, xromatinning strukturaviy tashkil etilishida ishtirok etadilar, bu esa yadroda DNKning zich qadoqlanishiga imkon beradi.
H2A, H2B, H3 va H4 gistonlarining har biri ikkita molekula DNKning 146-bp segmentiga o'ralgan oktamerni hosil qiladi, bu oqsil strukturasi tepasida spiralning 1,8 burilishini hosil qiladi. Bu 7 nm diametrli zarracha nukleosoma deb ataladi. Giston oktameri bilan bevosita aloqada bo'lmagan DNKning bir qismi (bog'lovchi DNK) giston H1 bilan o'zaro ta'sir qiladi.
Giston bo'lmagan oqsillar guruhi juda xilma-xil bo'lib, strukturaviy yadro oqsillarini, ko'plab fermentlarni va DNKning ma'lum bo'limlari bilan bog'liq bo'lgan transkripsiya omillarini va gen ekspressiyasini va boshqa jarayonlarni tartibga solishni o'z ichiga oladi.
Oktamerdagi gistonlar nukleosomadan chiqib turadigan 20 ta aminokislotadan iborat mobil N-terminal fragmentiga ("quyruq") ega bo'lib, xromatin tuzilishini saqlash va gen ekspressiyasini nazorat qilish uchun muhimdir. Masalan, xromosomalarning hosil bo'lishi (kondensatsiyasi) gistonlarning fosforlanishi bilan, transkripsiyaning kuchayishi esa ulardagi lizin qoldiqlarining atsetillanishi bilan bog'liq. Tartibga solish mexanizmining tafsilotlari to'liq tushunilmagan.
Nukleosoma - bu DNK va bitta giston H1 molekulasining to'rt juft giston oqsillari H2A, H2B, H3 va H4 to'plamidan tashkil topgan xromatin bo'linmasi. Histon H1 ikkita nukleosoma o'rtasidagi bog'lovchi DNK bilan bog'lanadi.
Nukleosoma xromatinning elementar qadoqlash birligidir. U sakkizta nukleosoma gistonining (giston oktameri) o'ziga xos kompleksiga o'ralgan DNK qo'sh spiralidan iborat. Nukleosoma diametri taxminan 11 nm bo'lgan disk shaklidagi zarracha bo'lib, nukleosoma gistonlarining har birining ikkita nusxasini (H2A, H2B, H3, H4) o'z ichiga oladi. Giston oktameri oqsil yadrosini hosil qiladi, uning atrofida ikki zanjirli DNK ikki marta o'raladi (har bir giston oktameriga 146 ta DNK asos jufti).
Fibrillalarni tashkil etuvchi nukleosomalar DNK molekulasi bo'ylab bir-biridan 10-20 nm masofada ko'proq yoki kamroq tekis joylashgan.
Eukariotlarda xromosomalarning qadoqlanish darajalari. Xromatin kondensatsiyasi.
Shunday qilib, DNK qadoqlash darajalari quyidagicha:
1) Nukleosomali (8 molekula giston oqsillari atrofida ikki zanjirli DNKning 2,5 burilishi).
2) Supernukleosoma - xromatin spiral (xromonema).
3) Xromatid - spirallashgan xromonema.
4) Xromosoma - DNK sperializatsiyasining to'rtinchi darajasi.
Interfaza yadrosida xromosomalar dekondensatsiyalanadi va xromatin bilan ifodalanadi. Genlarni o'z ichiga olgan o'ralgan joy euxromatin (bo'sh, tolali kromatin) deb ataladi. Bu transkripsiya uchun zaruriy shartdir. Bo'linishlar orasidagi uyqusizlik davrida ma'lum xromosoma hududlari va butun xromosomalar ixcham bo'lib qoladi.
Bu o'ralgan, juda bo'yalgan joylar geteroxromatin deb ataladi. Ular transkripsiyaviy jihatdan faol emas. Fakultativ va konstitutsiyaviy geteroxromatin mavjud.
Fakultativ heteroxromatin informativdir, chunki genlarni o'z ichiga oladi va evromatinga aylanishi mumkin. Ikki gomologik xromosomadan biri geteroxromatik bo'lishi mumkin. Konstitutsiyaviy geteroxromatin har doim geteroxromatik, shakllanmaydigan (genlarni o'z ichiga olmaydi) va shuning uchun transkripsiyada doimo faol emas.
Xromosoma DNK 108 dan ortiq tayanch juftlardan iborat bo'lib, ulardan informatsion bloklar - chiziqli joylashgan genlar hosil bo'ladi. Ular DNKning 25% ni tashkil qiladi. Gen - bu polipeptidlar yoki barcha RNK sintezi uchun ma'lumotni o'z ichiga olgan DNKning funktsional birligi. Genlar o'rtasida spacerlar - turli uzunlikdagi informatsion bo'lmagan DNK segmentlari mavjud. Ortiqcha genlar ko'p sonli - 104 ta bir xil nusxalar bilan ifodalanadi. Masalan, t-RNK, r-RNK va gistonlar uchun genlar. DNKda bir xil nukleotidlarning ketma-ketligi uchraydi. Ular o'rtacha takrorlanadigan yoki juda takrorlanadigan ketma-ketliklar bo'lishi mumkin. O'rtacha takrorlanuvchi ketma-ketliklar 102 - 104 takroriy takrorlanish bilan 300 ta nukleotid juftiga etadi va ko'pincha spacerlarni, ortiqcha genlarni ifodalaydi.
Ko'p takrorlanadigan ketma-ketliklar (105 - 106) konstitutsiyaviy heterokromatinni hosil qiladi. Barcha xromatinning taxminan 75% transkripsiyada ishtirok etmaydi, u juda takrorlanadigan ketma-ketliklardan va transkripsiyalanmagan oraliqlardan iborat.
Xromosoma preparatlarini tayyorlash. Kolxisindan foydalanish. Gipotoniya, fiksatsiya va bo'yash.
In vivo va in vitro turli to'qimalar hujayralarining proliferativ faollik darajasiga qarab xromosoma preparatlarini olishning bevosita va bilvosita usullari farqlanadi.
1) To'g'ridan-to'g'ri usullar mitotik faolligi yuqori bo'lgan to'qimalarni (suyak iligi, xorion va yo'ldosh, limfa tugunlari hujayralari, rivojlanishning dastlabki bosqichidagi embrion to'qimalarni) o'rganishda qo'llaniladi. Xromosoma preparatlari maxsus ishlovdan so'ng to'g'ridan-to'g'ri yangi olingan materialdan tayyorlanadi.
2) Bilvosita usullarga har qanday to'qimadan turli vaqtlar davomida oldindan o'stirilgandan so'ng xromosoma preparatlarini olish kiradi.
Xromosoma preparatlarini tayyorlashning to'g'ridan-to'g'ri va bilvosita usullarining ko'plab modifikatsiyalari mavjud, ammo metafaza plitalarini olishning asosiy bosqichlari o'zgarishsiz qolmoqda:
1. Kolxitsinni qo'llash (kolsemid) - metafaza bosqichida hujayra bo'linishini to'xtatuvchi mitotik shpindel shakllanishining inhibitori.
2. Kaliy yoki natriy tuzlari eritmalari yordamida gipotonik zarba, hujayralar ichidagi va tashqarisidagi osmotik bosimning farqi tufayli ularning shishishi va xromosomalararo bog'lanishlarning uzilishiga olib keladi. Ushbu protsedura xromosomalarning bir-biridan ajralishiga olib keladi va ularning metafaza plitalarida ko'proq tarqalishiga yordam beradi.
3. Muzlik sirka kislotasi va etanol (metanol) yordamida 3:1 nisbatda hujayralarni fiksatsiya qilish (Karnoy fiksatori), bu xromosoma tuzilishini saqlashga yordam beradi.
4. Hujayra suspenziyasini shisha slaydlarga tushirish.
5. Xromosoma preparatlarini bo'yash.
Xromosomadagi ko'ndalang belgilar (chiziqlar, tasmalar) majmuasini aniqlash uchun bir qancha bo'yash (bandlash) usullari ishlab chiqilgan. Har bir xromosoma o'ziga xos tasmalar majmuasi bilan tavsiflanadi. Gomologik xromosomalar bir xil bo'yalgan, genlarning turli xil allel variantlari lokalizatsiya qilingan polimorf hududlar bundan mustasno. Allelik polimorfizm ko'plab genlarga xosdir va ko'pchilik populyatsiyalarda uchraydi. Polimorfizmlarni sitogenetik darajada aniqlash diagnostik ahamiyatga ega emas.
A. Q-bo'yash. Xromosomalarni differentsial bo'yashning birinchi usuli shved sitologi Kaspersson tomonidan ishlab chiqilgan bo'lib, u bu maqsadda floresan bo'yoq xinin xantalidan foydalangan. Flüoresan mikroskop ostida xromosomalarda - Q-segmentlarida teng bo'lmagan floresans intensivligi bo'lgan joylar ko'rinadi. Usul Y xromosomalarini o'rganish uchun eng mos keladi va shuning uchun genetik jinsni tezda aniqlash, X va Y xromosomalari yoki Y xromosomalari va autosomalar orasidagi translokatsiyalarni (hududlar almashinuvini) aniqlash, shuningdek, ko'p sonli xromosomalarni ko'rish uchun ishlatiladi. jinsiy xromosoma mozaisizmi bo'lgan bemorda Y xromosomasi bo'lgan hujayralar kloniga ega yoki yo'qligini aniqlash zarur bo'lganda hujayralar.
B. G-bo'yash. Ko'pincha tripsin yordamida keng ko'lamli dastlabki ishlov berishdan so'ng, xromosomalar Giemsa bo'yog'i bilan bo'yaladi. Yorug'lik mikroskopi ostida xromosomalar - G-segmentlarida yorug'lik va quyuq chiziqlar ko'rinadi. Q segmentlarining joylashuvi G segmentlarining joylashishiga to'g'ri kelsa-da, G bo'yash sezgirroq ekanligini isbotladi va sitogenetik tahlilning standart usuli sifatida Q bo'yash o'rnini egalladi. G-bo'yash kichik aberatsiyalar va marker xromosomalarni (oddiy homolog xromosomalardan farqli ravishda segmentlangan) aniqlash uchun eng yaxshisidir.
B. R-bo'yash G-bo'yashga qarama-qarshi rasm beradi. Odatda Giemsa bo'yog'i yoki akridin apelsin floresan bo'yoq ishlatiladi. Bu usul opa-singillar xromatidlari yoki homolog xromosomalarning homolog G- yoki Q-manfiy hududlarini bo'yashdagi farqlarni ochib beradi.
G. C-bo'yash xromosomalarning sentromerik mintaqalarini (bu hududlarda konstitutsiyaviy geteroxromatin mavjud) va Y xromosomasining o'zgaruvchan, yorqin lyuminestsent distal qismini tahlil qilish uchun ishlatiladi.
E. T-bo'yash xromosomalarning telomer mintaqalarini tahlil qilish uchun ishlatiladi. Ushbu usul, shuningdek, yadroviy organizator hududlarni kumush nitrat bilan bo'yash (AgNOR bo'yash) standart xromosoma bo'yash orqali olingan natijalarni aniqlashtirish uchun ishlatiladi.
rRNK va tRNK prekursorlarining sintezi ire-mRNK sinteziga o'xshaydi. Ribosomal RNKning birlamchi transkriptida intronlar bo'lmaydi va o'ziga xos RNazalar ta'sirida u yorilib, 28S-, 18S- va 5,8S-rRNK hosil qiladi; 5S-pRNK RNK polimeraza III ishtirokida sintezlanadi.
rRNK va tRNK.
Birlamchi tRNK transkriptlari ham qisman gidroliz orqali etuk shakllarga aylanadi.
RNKning barcha turlari oqsil biosintezida ishtirok etadi, ammo bu jarayonda ularning vazifalari har xil. Oqsillarning birlamchi tuzilishini belgilovchi matritsaning roli messenjer RNK (mRNK) tomonidan amalga oshiriladi. Translatsiya mexanizmlarini o'rganish uchun hujayrasiz oqsil biosintezi tizimlaridan foydalanish muhim ahamiyatga ega. Agar to'qimalarning gomogenatlari aminokislotalar aralashmasi bilan inkubatsiya qilinsa, ulardan kamida bittasi etiketli bo'lsa, protein biosintezini oqsillarga yorliq kiritish orqali aniqlash mumkin. Sintezlanayotgan oqsilning birlamchi tuzilishi tizimga qo'shilgan mRNKning birlamchi tuzilishi bilan belgilanadi. Hujayrasiz tizim globin mRNK dan iborat bo'lsa (uni retikulotsitlardan ajratib olish mumkin), agar albumin gepatotsitlardan ajratilgan albumin mRNK bilan sintez qilinsa, globin (a- va 3-globin zanjirlari) sintezlanadi;
14. Replikatsiya ma’nosi:
a) jarayon proeukariotlarda hujayra boʻlinishining barcha turlariga asos boʻlgan muhim molekulyar mexanizmdir, b) bir hujayrali va koʻp hujayrali organizmlarning barcha turlarini koʻpaytirishni taʼminlaydi;
c) hujayraning doimiyligini saqlaydi
fiziologik regeneratsiya natijasida organlar, to'qimalar va tananing tarkibi
d) muayyan shaxslarning uzoq muddat yashashini ta'minlaydi;
e) organizmlar turlarining uzoq muddatli mavjudligini ta'minlaydi;
f) jarayon axborotning aniq ikki baravar ko'payishiga yordam beradi;
g) replikatsiya jarayonida patologik o'zgarishlar rivojlanishi bilan oqsil sintezining buzilishiga olib keladigan xatolar (mutatsiyalar) mumkin.
DNK molekulasining hujayra bo'linishidan oldin o'zini ko'paytirishning noyob xususiyati replikatsiya deb ataladi.
Irsiy ma'lumot tashuvchisi sifatida mahalliy DNKning o'ziga xos xususiyatlari:
1) replikatsiya - yangi zanjirlarning shakllanishi bir-birini to'ldiradi;
2) o'z-o'zini tuzatish - DNK polimeraza noto'g'ri replikatsiya qilingan bo'limlarni (10-6) ajratib oladi;
3) reparatsiya - tiklash;
Bu jarayonlar hujayrada maxsus fermentlar ishtirokida sodir bo'ladi.
Ta'mirlash tizimi qanday ishlaydi Qayta tiklash mexanizmlarini va bu qobiliyatning mavjudligini aniqlashga imkon bergan tajribalar bir hujayrali organizmlar yordamida amalga oshirildi. Ammo ta'mirlash jarayonlari hayvonlar va odamlarning tirik hujayralariga xosdir. Ba'zi odamlar xeroderma pigmentosumdan aziyat chekmoqda. Ushbu kasallik hujayralarning zararlangan DNKni qayta sintez qilish qobiliyatining etishmasligidan kelib chiqadi. Kseroderma meros qilib olinadi. Qayta tiklash tizimi nimadan iborat? Ta'mirlash jarayonini qo'llab-quvvatlaydigan to'rtta ferment DNK helikaz, -eksonukleaza, -polimeraza va -ligazadir. Ushbu birikmalarning birinchisi dezoksiribonuklein kislota molekulasi zanjiridagi zararni aniqlashga qodir. U nafaqat taniydi, balki molekulaning o'zgartirilgan segmentini olib tashlash uchun zanjirni to'g'ri joyda kesib tashlaydi. Yo'q qilishning o'zi DNK eksonukleaza yordamida amalga oshiriladi. Keyinchalik, shikastlangan qismni to'liq almashtirish uchun aminokislotalardan dezoksiribonuklein kislota molekulasining yangi bo'limi sintezlanadi. Xo'sh, bu eng murakkab biologik protseduraning yakuniy akkordi DNK ligaza fermenti yordamida amalga oshiriladi. U sintez qilingan joyni shikastlangan molekulaga biriktirish uchun javobgardir. To'rtta ferment ham o'z vazifasini bajarib bo'lgach, DNK molekulasi butunlay yangilanadi va barcha zararlar o'tmishda qoladi. Tirik hujayra ichidagi mexanizmlar shunday uyg'un ishlaydi.
Tasnifi Hozirgi vaqtda olimlar reparatsiya tizimlarining quyidagi turlarini ajratadilar. Ular turli omillarga qarab faollashadi. Bularga quyidagilar kiradi: Qayta faollashtirish. Rekombinatsiyani tiklash. Heteroduplex ta'mirlash. Eksizyonni tuzatish. DNK molekulalarining gomologik bo'lmagan uchlarining birlashishi. Barcha bir hujayrali organizmlar kamida uchta ferment tizimiga ega. Ularning har biri tiklanish jarayonini amalga oshirish qobiliyatiga ega. Ushbu tizimlarga quyidagilar kiradi: to'g'ridan-to'g'ri, eksizyon va post-replikativ. Prokaryotlar DNK ta'mirlashning ushbu uch turiga ega. Eukariotlarga kelsak, ularning ixtiyorida Miss-mathe va Sos-repair deb nomlangan qo'shimcha mexanizmlar mavjud. Biologiya hujayralarning genetik materialini o'z-o'zini davolashning barcha turlarini batafsil o'rganib chiqdi.
15. Genetik kod - barcha tirik organizmlarga xos bo'lgan nukleotidlar ketma-ketligi yordamida oqsillarning aminokislotalar ketma-ketligini kodlash usuli. Protein molekulasidagi aminokislotalar ketma-ketligi DNK molekulasidagi nukleotidlar ketma-ketligi sifatida shifrlangan va deyiladi. genetik kod. DNK molekulasining bitta oqsil sintezi uchun mas'ul bo'lgan qismi deyiladi genom.
DNK to'rtta nukleotiddan foydalanadi - adenin (A), guanin (G), sitozin (C), timin (T), rus adabiyotida A, G, C va T harflari bilan belgilanadi. Bu harflar alifboning alifbosini tashkil qiladi. genetik kod. RNK bir xil nukleotidlardan foydalanadi, timin bundan mustasno, u xuddi shunday nukleotid - urasil bilan almashtiriladi, u U harfi bilan belgilanadi (rus adabiyotida U). DNK va RNK molekulalarida nukleotidlar zanjir bo'lib joylashadi va shu tariqa genetik harflar ketma-ketligi olinadi.
Tabiatda oqsillarni qurish uchun 20 xil aminokislotalar ishlatiladi. Har bir oqsil qat'iy belgilangan ketma-ketlikda aminokislotalarning zanjiri yoki bir nechta zanjiridir. Bu ketma-ketlik oqsilning tuzilishini va shuning uchun uning barcha biologik xususiyatlarini aniqlaydi. Aminokislotalar to'plami ham deyarli barcha tirik organizmlar uchun universaldir.
Tirik hujayralarda genetik ma'lumotni amalga oshirish (ya'ni gen tomonidan kodlangan oqsil sintezi) ikkita matritsa jarayoni yordamida amalga oshiriladi: transkripsiya (ya'ni DNK matritsasida mRNK sintezi) va genetik kodni tarjima qilish. aminokislotalar ketma-ketligiga (mRNK matritsasida polipeptid zanjirining sintezi). 20 ta aminokislotalarni kodlash uchun ketma-ket uchta nukleotid, shuningdek, oqsil ketma-ketligining tugashini ko'rsatadigan to'xtash signali etarli. Uch nukleotidlar to'plamiga triplet deyiladi. Aminokislotalar va kodonlarga mos keladigan qabul qilingan qisqartmalar rasmda ko'rsatilgan.
Genetik kodning xususiyatlari
Triplet - kodning mazmunli birligi uchta nukleotidning (uchlik yoki kodon) birikmasidir.
Davomiylik - uchlik o'rtasida tinish belgilari yo'q, ya'ni ma'lumot uzluksiz o'qiladi.
Bir-biriga yopishmaslik - bir xil nukleotid bir vaqtning o'zida ikki yoki undan ortiq tripletlarning bir qismi bo'lishi mumkin emas. (Viruslar, mitoxondriyalar va bakteriyalardagi ba'zi bir-biriga o'xshash genlar uchun to'g'ri kelmaydi, ular bir nechta ramka almashinuvi oqsillarini kodlaydi.)
O'ziga xoslik - ma'lum bir kodon faqat bitta aminokislotaga to'g'ri keladi. (Xususiyat universal emas. Euplotes crassusdagi UGA kodoni ikkita aminokislota - sistein va selenotsisteinni kodlaydi)
Degeneratsiya (ortiqchalik) - bir xil aminokislotaga bir nechta kodon mos kelishi mumkin.
Umumjahonlik - genetik kod turli darajadagi murakkablikdagi organizmlarda bir xil ishlaydi - viruslardan tortib to odamlargacha (genetik muhandislik usullari bunga asoslanadi) (Shuningdek, bu xususiyatga bir qator istisnolar ham mavjud, "Variatsiyalar" jadvaliga qarang. ushbu maqoladagi standart genetik kod" bo'limi).
16.Biosintez sharoitlari
Protein biosintezi DNK molekulasidan genetik ma'lumotni talab qiladi; xabarchi RNK - bu ma'lumotni yadrodan sintez joyiga tashuvchi; ribosomalar - oqsil sintezining o'zi sodir bo'ladigan organellalar; sitoplazmadagi aminokislotalar to'plami; aminokislotalarni kodlaydigan RNKlarni o'tkazish va ularni ribosomalardagi sintez joyiga o'tkazish; ATP kodlash va biosintez jarayonini energiya bilan ta'minlaydigan moddadir.
Bosqichlar
Transkripsiya- yadroda sodir bo'ladigan DNK matritsasidagi barcha turdagi RNKning biosintezi jarayoni.
DNK molekulasining ma'lum bir qismi tushkunlikka tushadi, ikkita zanjir orasidagi vodorod aloqalari fermentlar ta'sirida yo'q qilinadi. Bir DNK zanjirida, shablonda bo'lgani kabi, RNK nusxasi komplementar printsipga muvofiq nukleotidlardan sintezlanadi. DNK bo'limiga qarab ribosoma, transport va xabarchi RNKlar shu tarzda sintezlanadi.
mRNK sintezidan so'ng u yadroni tark etadi va sitoplazmaga ribosomalarda oqsil sintezi joyiga yuboriladi.
Translyatsiya- ribosomalarda amalga oshiriladigan polipeptid zanjirlarini sintez qilish jarayoni, bu erda mRNK oqsilning birlamchi tuzilishi haqida ma'lumot uzatishda vositachi hisoblanadi.
Protein biosintezi bir qator reaksiyalardan iborat.
1. Aminokislotalarning faollashishi va kodlanishi. tRNK yonca bargi shakliga ega, uning markaziy halqasida ma'lum bir aminokislota va mRNKdagi kodon kodiga mos keladigan uchlik antikodon joylashgan. Har bir aminokislota ATP energiyasidan foydalangan holda tegishli tRNK bilan bog'lanadi. tRNK-aminokislotalar kompleksi hosil bo'lib, u ribosomalarga kiradi.
2. mRNK-ribosoma kompleksining shakllanishi. Sitoplazmadagi mRNK donador ER dagi ribosomalar bilan bog'langan.
3. Polipeptid zanjirining yig'ilishi. Aminokislotalar bilan tRNK, antikodon-kodon komplementarligi printsipiga ko'ra, mRNK bilan birlashadi va ribosomaga kiradi. Ribosomaning peptid markazida ikkita aminokislotalar o'rtasida peptid bog'i hosil bo'ladi va ajralib chiqqan tRNK ribosomani tark etadi. Bunday holda, mRNK har safar bitta tripletni oldinga siljitadi, yangi tRNK - aminokislota kiritadi va ribosomadan chiqarilgan tRNKni olib tashlaydi. Butun jarayon ATP energiyasi bilan ta'minlanadi. Bitta mRNK bir nechta ribosomalar bilan birlashib, bir vaqtning o'zida bitta oqsilning ko'plab molekulalari sintezlanadigan polisoma hosil qilishi mumkin. Sintez mRNKda bema'ni kodonlar (to'xtash kodlari) boshlanganda tugaydi. Ribosomalar mRNK dan ajratiladi va ulardan polipeptid zanjirlari chiqariladi. Butun sintez jarayoni granulyar endoplazmatik retikulumda sodir bo'lganligi sababli, hosil bo'lgan polipeptid zanjirlari ER kanalchalariga kiradi va u erda ular oxirgi tuzilishga ega bo'ladi va oqsil molekulalariga aylanadi.
Barcha sintez reaktsiyalari ATP energiyasini sarflash bilan maxsus fermentlar tomonidan katalizlanadi. Sintez tezligi juda yuqori va polipeptid uzunligiga bog'liq. Masalan, ichak tayoqchasi ribosomasida 300 ta aminokislotadan iborat oqsil taxminan 15-20 soniyada sintezlanadi.